
Comment le transport de matériaux est-il organisé au sein des cellules ? Pour de nombreuses molécules, il se fait dans des petits sacs, des vésicules, dont l’enveloppe est une membrane semblable à celle qui délimite la cellule. Par exemple, au niveau d’une synapse entre deux neurones, les messagers chimiques (les neurotransmetteurs) sont enfermés dans des vésicules qui circulent dans la cellule avant de fusionner avec la membrane cellulaire : de la sorte, le contenu est libéré à l’extérieur du neurone.
Les trois lauréats ont élucidé les principes moléculaires qui gouvernent ce trafic. L’Américain Randy Schekman, de l’Université de Californie, à Berkeley, a découvert un ensemble de gènes impliqués dans ce transport. L’Américain James Rothman, de l’Université Yale, a identifié les protéines grâce auxquelles les vésicules fusionnent avec les membranes cellulaires. Enfin, l’Allemand Thomas Südhof, de l’Université Stanford, a mis en évidence les signaux qui déclenchent cette fusion. Voyons en détail ce système d’import et d’export dans la cellule.
Dans une cellule, l’espace est compartimenté dans plusieurs organelles, des zones délimitées par des membranes : citons notamment l’appareil de Golgi et le réticulum endoplasmique(voir la figure). les diverses molécules produites (neurotransmetteurs, hormones, enzymes, cytokines) ne sont pas « libres » dans le cytoplasme, mais le plus souvent enfermées dans des vésicules qui se déplacent entre les différents compartiments et vers la membrane cellulaire, porte d’accès vers l’extérieur. Ces vésicules sont des sortes de cargos qui doivent délivrer leur contenu au bon moment et à la bonne place.
Dans les années 1970, R. Schekman étudie la levure Saccharomyces cerevisiae (la levure de boulanger) et en identifie quelques unes chez qui le système de transport est déficient : dans certains cas, les vésicules s’entassent sans jamais libérer leur cargaison. Des analyses génétiques mettent en lumière 23 gènes incriminés dans ces défauts. Ces gènes se répartissent en trois catégories, qui renvoient à l’appareil de Golgi, au réticulum endoplasmique et à la membrane cellulaire. R. Schekman montre alors une voie de transport entre le réticulum endoplasmique et l’appareil de Golgi.
La décennie suivant, J. Rothman met au jour les complexes protéiques qui permettent aux vésicules de s’arrimer et de fusionner avec la membrane cellulaire. Des protéines situées sur chacune des membranes s’associent à la façon d’une fermeture éclair. Seules certaines combinaisons de protéines étant autorisées, ce système autorise un adressage des vésicules, qui ne peuvent pas fusionner n’importe où. Ces mécanismes ont également cours à l’intérieur de la cellule entre les compartiments. La première protéine identifiée était NSF (N-Ethylmaleimide-sensitive factor). D’autres ont suivi, telles les protéines SNARE (voir la figure). Certains de gènes identifiés par R. Schekman chez la levure correspondaient aux protéines découvertes par J. Rothman chez les mammifères, montrant une origine évolutive ancienne de ces mécanismes.
T. Südhof s’intéressait à la communication entre cellules nerveuses. Au niveau de la synapse, les signaux sont des molécules qui sont libérées lors de la fusion de vésicules les contenant avec la membrane cellulaire. Les mécanismes sont ceux mis en évidence par R. Schekman et J. Rothman. Cependant, la fusion n’a lieu que lorsqu’un influx nerveux parvient à la synapse. On connaissait le rôle des ions calcium dans ce processus ; T. Südhof a découvert les protéines sensibles au calcium qui font le lien entre le signal électrique et la fusion des vésicules contenant les neurotransmetteurs.
À eux trois, les lauréats ont révélé les détails d’un système de transport cellulaire complexe contrôlé de façon très précise. Des dérèglements seraient en cause dans diverses maladies neurologiques, mais aussi dans le diabète et des troubles immunologiques.
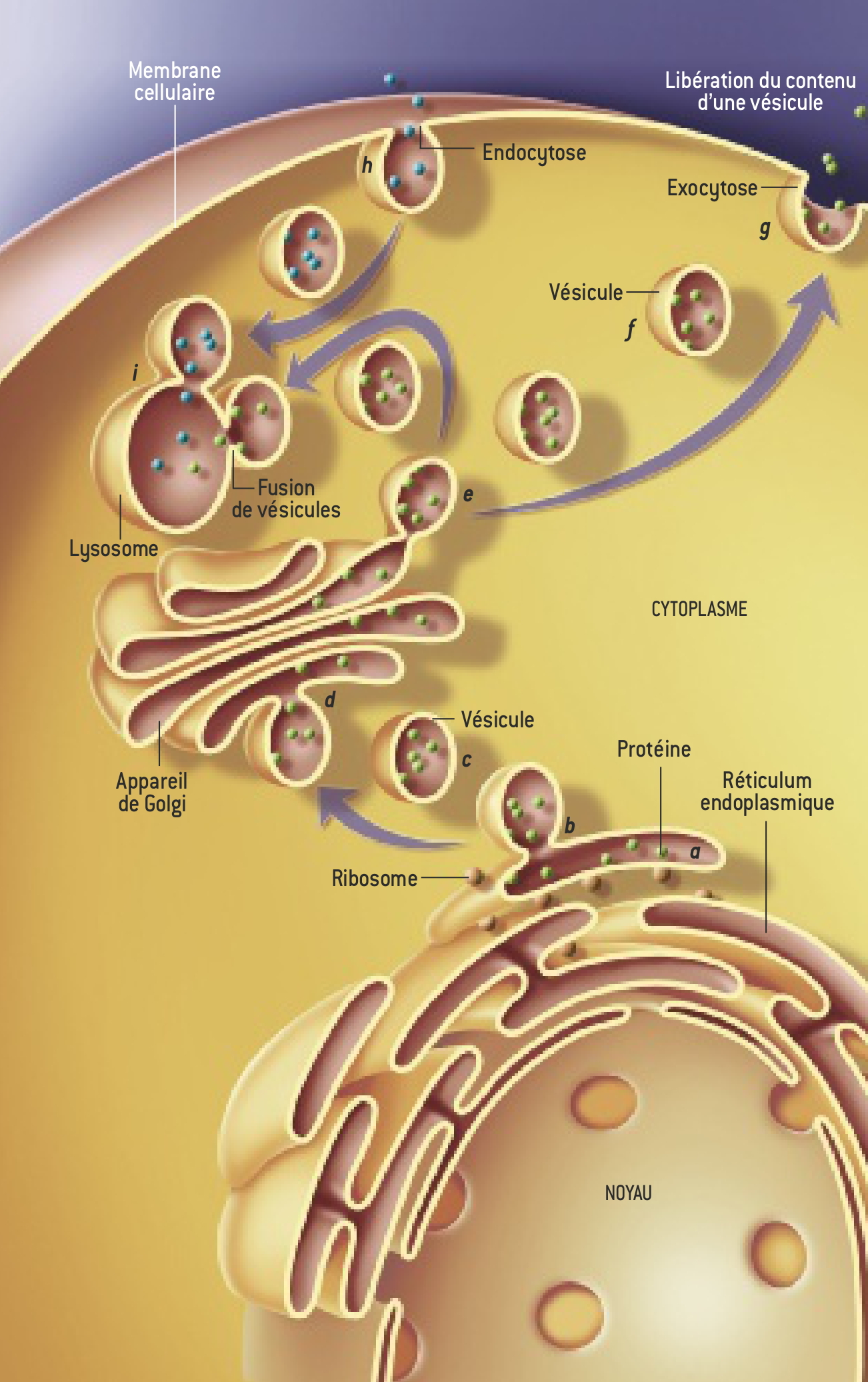
Le trafic intracellulaire. Les différents organites, c’est-à-dire les éléments de la cellule limités par une membrane lipidique, échangent des molécules grâce à des transporteurs, des vésicules elles aussi lipidiques. Par exemple, une protéine fabriquées dans le cytoplasme à la périphérie du noyau est incorporée dans le réseau de saccules du réticulum endoplasmique (a). Elle quitte cet organite dans une vésicule qui bourgeonne (b) à partir de la membrane du réticulum, s’en détache (c) et fusionne (d) avec celle de l’appareil de Golgi. Elle sera libérée hors de la cellule par la même série d’étapes (de e à g), la vésicule fusionnant cette fois avec la membrane cellulaire : ce mécanisme d’excrétion est l’exocytose. L’endocyotse (h) est un processus inverse. Dans le cytoplasme, plusieurs vésicules peuvent aussi fusionner (i).

La fusion des membranes, par exemple celle d’une vésicule et celle d’une cellule, résulte de l’association de protéines SNARE (en vert et en violet) portées par les deux membranes (a). Ces protéines s’enroulent en hélice et rapprochent ainsi les deux membranes dont les deux feuillets en contact fusionnent d’abord (b). Cette forme intermédiaire est dite en « trognon de pomme ». Puis les deux compartiments sont réunis via un pore (c) qui s’élargit progressivement (d) : la livraison est effectuée.
